Objectives: People living with HIV (PLWH) with low CD4 T-cell counts may be at a higher risk for severe coronavirus disease 2019 (COVID-19) outcomes and in need of efficient vaccination. The World Health Organization (WHO) now recommends prioritizing PLHIV for COVID-19 vaccination. Data on immune responses after messenger RNA (mRNA) vaccination in PLHIV in relation to CD4 counts are scarce. We aimed at assessing the humoral immune response in PLHIV after mRNA vaccination against COVID-19.
Methods: We examined a cohort of PLHIV after prime (n = 88) and boost (n = 52) vaccination with BNT162b2. We assessed levels of anti-severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spike (S) protein-specific immunoglobulin G (IgG)/IgA and circulating neutralizing antibodies in plasma and correlated results to the cellular immune status. BNT162b2-vaccinated health care workers served as controls.
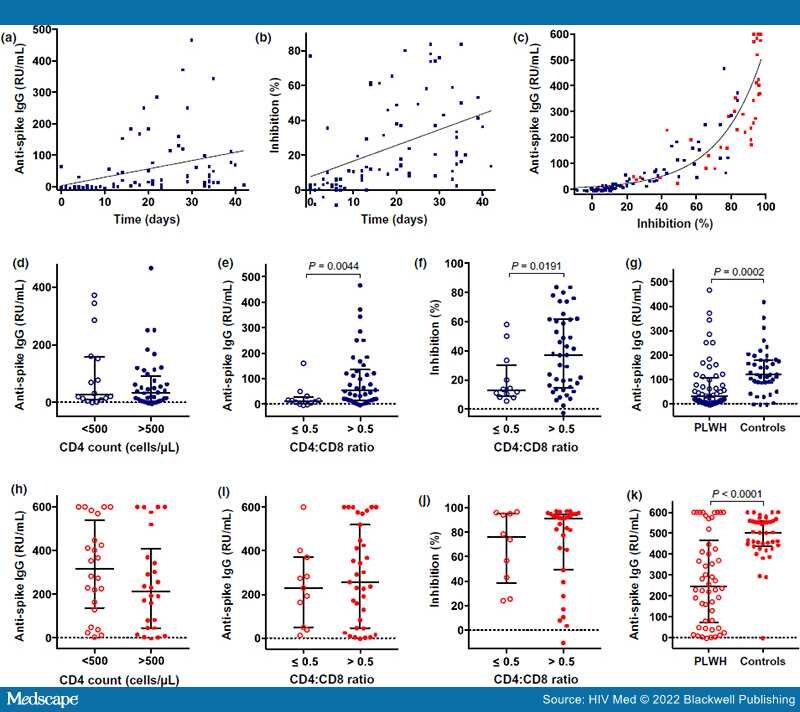
Results:All PLWH had a viral load of ≤ 200 HIV-1 RNA copies/mL and 96.5% had a viral load of < 50 copies/mL. Anti-S IgG and neutralizing antibody responses after BNT162b2 priming were significantly lower in PLHIV having a CD4:CD8 T-cell ratio of < 0.5. However, we observed robust humoral immunity in the majority of PLWH receiving antiretroviral therapy (ART) irrespective of CD4 T-cell nadir, current CD4 count or CD4:CD8 ratio after full BNT162b2 vaccination. Nevertheless, HIV-negative controls produced significantly higher mean anti-S IgG concentrations with less variability.